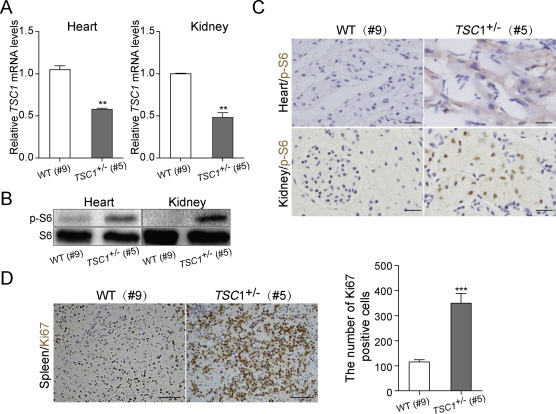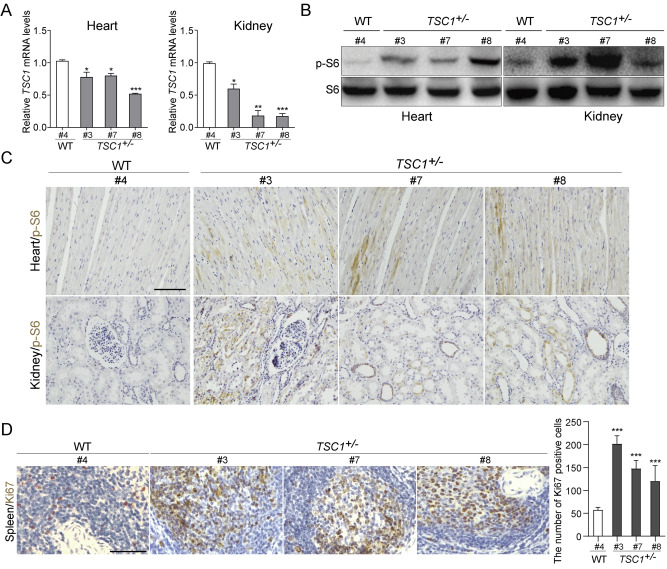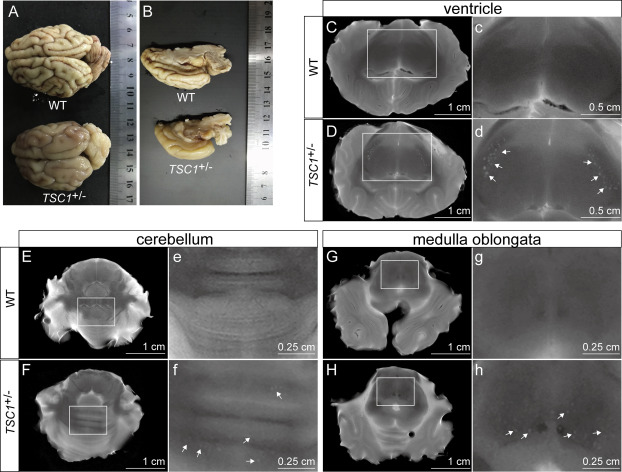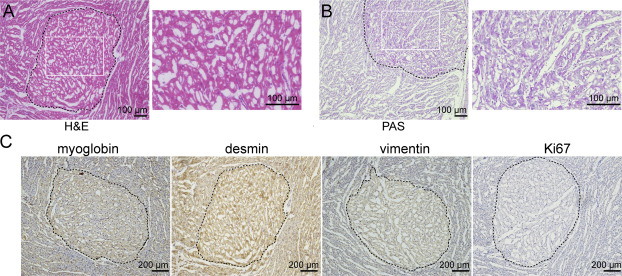
| [1] |
Au, K.S., Williams, A.T., Roach, E.S., Batchelor, L., Sparagana, S.P., Delgado, M.R., Wheless, J.W., Baumgartner, J.E., Roa, B.B., Wilson, C.M., Smith-Knuppel, T.K., Cheung, M.Y., Whittemore, V.H., King, T.M., Northrup, H., 2007. Genotype/phenotype correlation in 325 individuals referred for a diagnosis of tuberous sclerosis complex in the United States. Genet. Med. 9, 88-100.
|
| [2] |
Aw, F., Goyer, I., Raboisson, M.J., Boutin, C., Major, P., Dahdah, N., 2017. Accelerated cardiac rhabdomyoma regression with everolimus in infants with tuberous sclerosis complex. Pediatr. Cardiol. 38, 394-400.
|
| [3] |
Ben-Sahra, I., Manning, B.D., 2017. mTORC1 signaling and the metabolic control of cell growth. Curr. Opin. Cell Biol. 45, 72-82.
|
| [4] |
Cao, J., Tyburczy, M.E., Moss, J., Darling, T.N., Widlund, H.R., Kwiatkowski, D.J., 2017. Tuberous sclerosis complex inactivation disrupts melanogenesis via mTORC1 activation. J. Clin. Invest. 127, 349-364.
|
| [5] |
Chen, F., Wang, Y., Yuan, Y., Zhang, W., Ren, Z., Jin, Y., Liu, X., Xiong, Q., Chen, Q., Zhang, M., Li, X., Zhao, L., Li, Z., Wu, Z., Zhang, Y., Hu, F., Huang, J., Li, R., Dai, Y., 2015. Generation of B cell-deficient pigs by highly efficient CRISPR/Cas9-mediated gene targeting. J. Genet. Genomics 42, 437-444.
|
| [6] |
Chen, Q., Fang, B., Wang, Y., Li, C., Li, X., Wang, R., Xiong, Q., Zhang, L., Jin, Y., Zhang, M., Liu, X., Li, L., Mou, L., Li, R., Yang, H., Dai, Y., 2018. Overexpressing dominant-negative FGFR2-IIIb impedes lung branching morphogenesis in pigs. J. Genet. Genomics 45, 147-154.
|
| [7] |
Curatolo, P., Moavero, R., Roberto, D., Graziola, F., 2015. Genotype/phenotype correlations in tuberous sclerosis complex. Semin. Pediatr. Neurol. 22, 259-273.
|
| [8] |
DiMario, F.J., Jr., Sahin, M., Ebrahimi-Fakhari, D., 2015. Tuberous sclerosis complex. Pediatr. Clin. North Am. 62, 633-648.
|
| [9] |
Fang, B., Ren, X., Wang, Y., Li, Z., Zhao, L., Zhang, M., Li, C., Zhang, Z., Chen, L., Li, X., Liu, J., Xiong, Q., Zhang, L., Jin, Y., Liu, X., Li, L., Wei, H., Yang, H., Li, R., Dai, Y., 2018. Apolipoprotein E deficiency accelerates atherosclerosis development in miniature pigs. Dis. Mod. Mech. 11, dmm036632.
|
| [10] |
Fu, D.A., Campbell-Thompson, M., 2017. Periodic acid-schiff staining with diastase. Methods Mol. Bio. 1639, 145-149.
|
| [11] |
Guo, Y., Dreier, J.R., Cao, J., Du, H., Granter, S.R., Kwiatkowski, D.J., 2016. Analysis of a mouse skin model of tuberous sclerosis complex. PLoS One 11, e0167384.
|
| [12] |
Hai, T., Guo, W., Yao, J., Cao, C., Luo, A., Qi, M., Wang, X., Wang, X., Huang, J., Zhang, Y., Zhang, H., Wang, D., Shang, H., Hong, Q., Zhang, R., Jia, Q., Zheng, Q., Qin, G., Li, Y., Zhang, T., Jin, W., Chen, Z.Y., Wang, H., Zhou, Q., Meng, A., Wei, H., Yang, S., Zhao, J., 2017. Creation of miniature pig model of human Waardenburg syndrome type 2A by ENU mutagenesis. Hum. Genet. 136, 1463-1475.
|
| [13] |
Islam, M.P., Roach, E.S., 2015. Tuberous sclerosis complex. Handb. Clin. Neurol. 132, 97-109.
|
| [14] |
Kobayashi, T., Minowa, O., Kuno, J., Mitani, H., Hino, O., Noda, T., 1999. Renal carcinogenesis, hepatic hemangiomatosis, and embryonic lethality caused by a germline Tsc2 mutation in mice. Cancer Res. 59, 1206-1211.
|
| [15] |
Kobayashi, T., Minowa, O., Sugitani, Y., Takai, S., Mitani, H., Kobayashi, E., Noda, T., Hino, O., 2001. A germ-line Tsc1 mutation causes tumor development and embryonic lethality that are similar, but not identical to, those caused by Tsc2 mutation in mice. Proc. Natl. Acad. Sci. U. S. A. 98, 8762-8767.
|
| [16] |
Krueger, D.A., Northrup, H., 2013. Tuberous sclerosis complex surveillance and management: recommendations of the 2012 International Tuberous Sclerosis Complex Consensus Conference. Pediatr. Neurol. 49, 255-265.
|
| [17] |
Kwiatkowski, D.J., 2010. Animal models of lymphangioleiomyomatosis (LAM) and tuberous sclerosis complex (TSC). Lymphat. Res. Biol. 8, 51-57.
|
| [18] |
Kwiatkowski, D.J., Zhang, H., Bandura, J.L., Heiberger, K.M., Glogauer, M., el-Hashemite, N., Onda, H., 2002. A mouse model of TSC1 reveals sex-dependent lethality from liver hemangiomas, and up-regulation of p70S6 kinase activity in Tsc1 null cells. Hum. Mol. Genet. 11, 525-534.
|
| [19] |
Lam, H.C., Nijmeh, J., Henske, E.P., 2017. New developments in the genetics and pathogenesis of tumours in tuberous sclerosis complex. J. Pathol. 241, 219-225.
|
| [20] |
Lu, D.S., Karas, P.J., Krueger, D.A., Weiner, H.L., 2018. Central nervous system manifestations of tuberous sclerosis complex. Am. J. Med. Genet. C. Semin. Med. Genet. 178, 291-298.
|
| [21] |
Martin, K.R., Zhou, W., Bowman, M.J., Shih, J., Au, K.S., Dittenhafer-Reed, K.E., Sisson, K.A., Koeman, J., Weisenberger, D.J., Cottingham, S.L., DeRoos, S.T., Devinsky, O., Winn, M.E., Cherniack, A.D., Shen, H., Northrup, H., Krueger, D.A., MacKeigan, J.P., 2017. The genomic landscape of tuberous sclerosis complex. Nat. Commun. 8, 15816.
|
| [22] |
Onda, H., Lueck, A., Marks, P.W., Warren, H.B., Kwiatkowski, D.J., 1999. Tsc2(+/-) mice develop tumors in multiple sites that express gelsolin and are influenced by genetic background. J. Clin. Invest. 104, 687-695.
|
| [23] |
Pfaffl, M.W., 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29, e45.
|
| [24] |
Portocarrero, L.K.L., Quental, K.N., Samorano, L.P., Oliveira, Z.N.P., Rivitti-Machado, M., 2018. Tuberous sclerosis complex: review based on new diagnostic criteria. An. Bras. Dermatol. 93, 323-331.
|
| [25] |
Randle, S.C., 2017. Tuberous Sclerosis Complex: A Review. Pediatr. Ann. 46, e166-e171.
|
| [26] |
Rubilar, C., Lopez, F., Troncoso, M., Barrios, A., Herrera, L., 2017. [Clinical and genetic study patients with tuberous sclerosis complex]. Rev. Chil. Pediatr. 88, 41-49.
|
| [27] |
Salussolia, C.L., Klonowska, K., Kwiatkowski, D.J., Sahin, M., 2019. Genetic etiologies, diagnosis, and treatment of tuberous sclerosis complex. Annu. Rev. Genomics Hum. Genet. 20, 217-240.
|
| [28] |
Saxton, R.A., Sabatini, D.M., 2017. mTOR signaling in growth, metabolism, and disease. Cell 168, 960-976.
|
| [29] |
Shen, Q., Shen, J., Qiao, Z., Yao, Q., Huang, G., Hu, X., 2015. Cardiac rhabdomyomas associated with tuberous sclerosis complex in children. From presentation to outcome. Herz 40, 675-678.
|
| [30] |
Uzun, O., Wilson, D.G., Vujanic, G.M., Parsons, J.M., De Giovanni, J.V., 2007. Cardiac tumours in children. Orphanet J. Rare Dis. 2, 11.
|
| [31] |
van Slegtenhorst, M., Verhoef, S., Tempelaars, A., Bakker, L., Wang, Q., Wessels, M., Bakker, R., Nellist, M., Lindhout, D., Halley, D., van den Ouweland, A., 1999. Mutational spectrum of the TSC1 gene in a cohort of 225 tuberous sclerosis complex patients: no evidence for genotype-phenotype correlation. J. Med. Genet. 36, 285-289.
|
| [32] |
Yan, S., Tu, Z., Liu, Z., Fan, N., Yang, H., Yang, S., Yang, W., Zhao, Y., Ouyang, Z., Lai, C., Yang, H., Li, L., Liu, Q., Shi, H., Xu, G., Zhao, H., Wei, H., Pei, Z., Li, S., Lai, L., Li, X.J., 2018. A Huntingtin knockin pig model recapitulates features of selective neurodegeneration in Huntington's disease. Cell 173, 989-1002.e13.
|
| [33] |
Yao, J., Zeng, H., Zhang, M., Wei, Q., Wang, Y., Yang, H., Lu, Y., Li, R., Xiong, Q., Zhang, L., Chen, Z., Xing, G., Cao, X., Dai, Y., 2019. OSBPL2-disrupted pigs recapitulate dual features of human hearing loss and hypercholesterolaemia. J. Genet. Genomics 46, 379-387.
|
| [34] |
Yu, Z., Zhang, X., Guo, H., Bai, Y., 2014. A novel TSC2 mutation in a Chinese family with tuberous sclerosis complex. J. Genet. 93, 169-172.
|
| [35] |
Zhang, R., Wang, Y., Chen, L., Wang, R., Li, C., Li, X., Fang, B., Ren, X., Ruan, M., Liu, J., Xiong, Q., Zhang, L., Jin, Y., Zhang, M., Liu, X., Li, L., Chen, Q., Pan, D., Li, R., Cooper, D.K.C., Yang, H., Dai, Y., 2018. Reducing immunoreactivity of porcine bioprosthetic heart valves by genetically-deleting three major glycan antigens, GGTA1/β4GalNT2/CMAH. Acta Biomater. 72, 196-205.
|
Figures (5)





Supported by: Beijing Renhe Information Technology Co. Ltd




 DownLoad:
DownLoad: