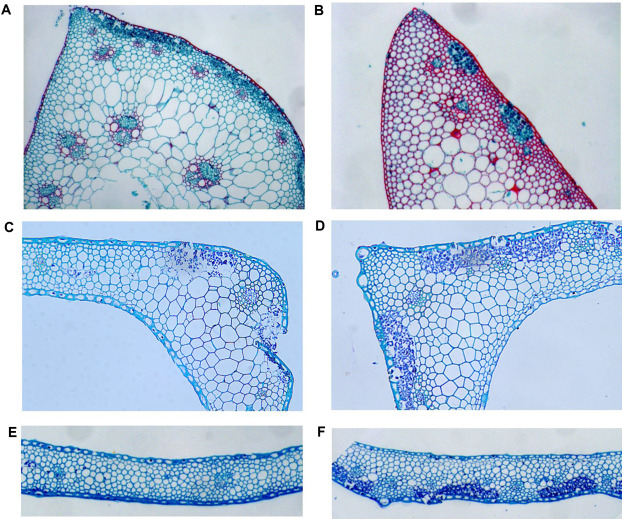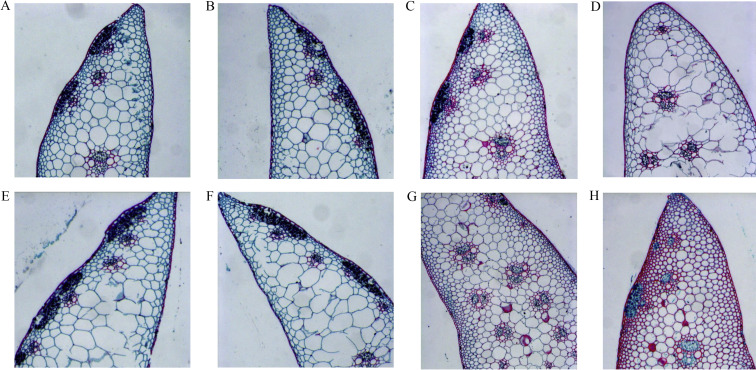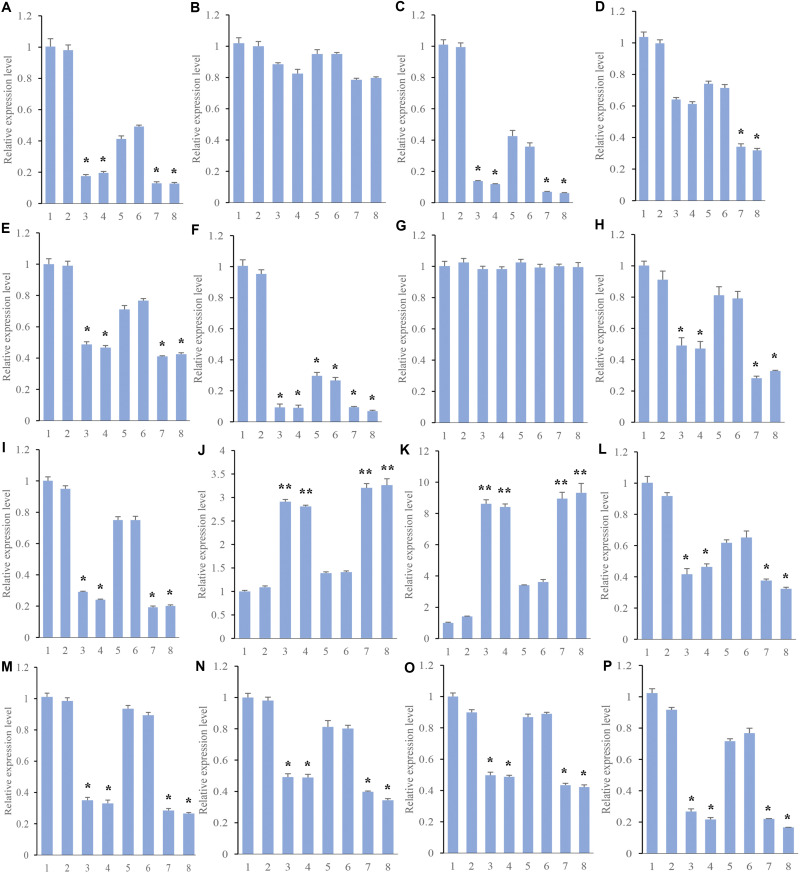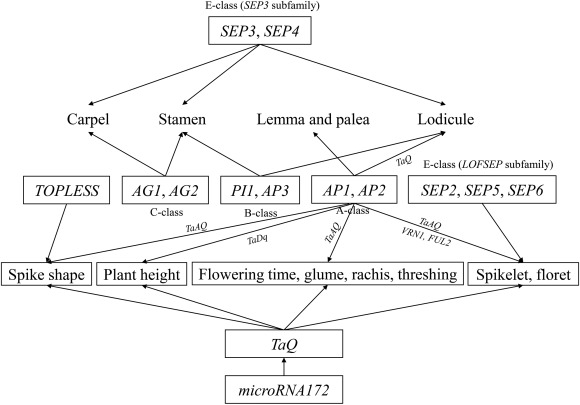
| [1] |
Abe, F., Haque, E., Hisano, H., Tanaka, T., Kamiya, Y., Mikami, M., Kawaura, K., Endo, M., Onishi, K., Hayashi, T., Sato, K., 2019. Genome-edited triple-recessive mutation alters seed dormancy in wheat. Cell Rep.. 28, 1362-1369.
|
| [2] |
Aukerman, M.J., Sakai, H., 2003. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like target genes. Plant Cell 15, 2730-2741.
|
| [3] |
Brown, R.H., Bregitzer, P., 2011. A Ds insertional mutant of a barley miR172 gene results in indeterminate spikelet development. Crop Sci. 51, 1664-1672.
|
| [4] |
Causier, B., Ashworth, M., Guo, W., Davies, B., 2012. The TOPLESS interactome: a framework for gene repression in Arabidopsis. Plant Physiol.. 158, 423-438.
|
| [5] |
Chuck, G., Meeley, R., Hake, S., 2008. Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1. Development 135, 3013-3019.
|
| [6] |
Chuck, G., Meeley, R., Irish, E., Sakai, H., Hake, S., 2007. The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1. Nat. Genet. 39, 1517-1521.
|
| [7] |
Cui, R., Han, J., Zhao, S., Su, K., Wu, F., Du X, Xu, Q., Chong, K., Theissen, G., Meng, Z., 2010. Functional conservation and diversification of class E floral homeotic genes in rice (Oryza sativa). Plant J.. 61, 767-781.
|
| [8] |
Debernardi, J.M., Greenwood, J.R., Finnegan, E.J., Jernstedt, J., Dubcovsky, J., 2020. APETALA2-like genes AP2L2 and Q specify lemma identity and axillary floral meristem development in wheat. Plant J.. 101, 171-187.
|
| [9] |
Debernardi, J.M., Lin, H., Chuck, G., Faris, J.D., Dubcovsky, J., 2017. microRNA172 plays a crucial role in wheat spike morphogenesis and grain threshability. Development 144, 1966-1975.
|
| [10] |
Ditta, G., Stanfield, S., Corbin, D., Helinski, D.R., 1980. Broad host range DNA cloning system for gram-negative bacteria: construction of a gene bank of Rhizobium meliloti. Proc. Natl. Acad. Sci. U. S. A. 77, 7347-7351.
|
| [11] |
Drews, G.N., Bowman, J.L., Meyerowitz, E.M., 1991. Negative regulation of the Arabidopsis homeotic gene AGAMOUS by the APETALA2 product. Cell 65, 991-1002.
|
| [12] |
Faris, J.D., Simons, K.J., Zhang, Z., Gill, B.S., 2005. The wheat super domestication gene Q. Front. Wheat Biosci. 100, 129-148.
|
| [13] |
Hong, L.L., Qian, Q., Zhu, K.M., Tang, D., Huang, Z.J., Gao, L., Li, M., Gu, M.H., Cheng, Z.K., 2010. ELE restrains empty glumes from developing into lemmas. J. Genet. Genomics 37, 101-115.
|
| [14] |
Howells, R.M., Craze, M., Bowden, S., Wallington, E.J., 2018. Efficient generation of stable, heritable gene edits in wheat using CRISPR/Cas9. BMC Plant Biol.. 18, 215.
|
| [15] |
10.1007/978-1-4939-1695-5_15 |
| [16] |
Jiang, Y.F., Chen, Q., Wang, Y., Guo, Z.R., Xu, B.J., Zhu, J., Zhang, Y.Z., Gong, X., Luo, C.H., Wu, W., Liu, C.H., Kong, L., Deng, M., Jiang, Q.T., Lan, X.J., Wang, J.R., Chen, G.Y., Zheng, Y.L., Wei, Y.M., Qi, P.F., 2019. Re-acquisition of the brittle rachis trait via a transposon insertion in domestication gene Q during wheat de-domestication. New Phytol.. 224, 961-973.
|
| [17] |
Jiao, R., Gao, C., 2016. The CRISPR/Cas9 genome editing revolution. J. Genet. Genomics 43, 227-228.
|
| [18] |
Jung, J.H., Seo, Y.H., Seo, P.J., Reyes, J.L., Yun, J., Chua, N.H., Park, C.M., 2007. The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis. Plant Cell 19, 2736-2748.
|
| [19] |
Kato, K., Sonokawa, R., Miura, H., Sawada, S., 2003. Dwarfing effect associated with the threshability gene Q on wheat chromosome 5A. Plant Breed. 122, 489-492.
|
| [20] |
Lee, D.Y., An, G., 2012. Two AP2 family genes, supernumerary bract (SNB) and Osindeterminate spikelet 1 (OsIDS1), synergistically control inflorescence architecture and floral meristem establishment in rice. Plant J.. 69, 445-461.
|
| [21] |
Liang, Z., Chen, K.L., Li, T.D., Zhang, Y., Wang, Y.P., Zhao, Q., Liu, J.X., Zhang, H.W., Liu, C.M., Ran, Y.D., Gao, C.X., 2017. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 8, 14261.
|
| [22] |
Liang, Z., Chen, K.L., Zhang, Y., Liu, J.X., Yin, K.Q., Qiu, J.L., Gao, C.X., 2018. Genome editing of bread wheat using biolistic delivery of CRISPR/Cas9 in vitro transcripts or ribonucleoproteins. Nat. Protoc. 13, 413-430.
|
| [23] |
Li, C., Lin, H., Chen, A., Lau, M., Jernstedt, J., Dubcovsky, J., 2019. Wheat VRN1 and FUL2 play critical and redundant roles in spikelet development and spike determinacy. Development 146, dev175398.
|
| [24] |
Liu, C., Zhong, Y., Qi, X., Chen, M., Liu, Z., Tian, X., Li, J., Jiao, Y., Wang, D., Wang, Y., Li, M., Xin, M., Liu, W., Jin, W., Chen, S., 2020a. Extension of the in vivo haploid induction system from maize to wheat. Plant Biotechnol. J. 18, 316-318.
|
| [25] |
Liu, H., Wang, K., Jia, Z., Gong, Q., Lin, Z., Du, L., Pei, X., Ye, X., 2020b. Efficient induction of haploid plants in wheat by editing of TaMTL using an optimized Agrobacterium-mediated CRISPR system. J. Exp. Bot. 71, 1337-1349.
|
| [26] |
Liu, P., Liu, J., Dong, H., Sun, J., 2018. Functional regulation of Q by microRNA172 and transcriptional co-repressor TOPLESS in controlling bread wheat spikelet density. Plant Biotechnol. J. 16, 495-506.
|
| [27] |
Ma, X., Zhang, Q., Zhu, Q., Liu, W., Chen, Y., Qiu, R., Wang, B., Yang, Z., Li, H., Lin, Y., Xie, Y., Shen, R., Chen, S., Wang, Z., Chen, Y., Guo, J., Chen, L., Zhao, X., Dong, Z., Liu, Y.G., 2015. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 8, 1274-1284.
|
| [28] |
Malcomber, S.T., Kellogg, E.A., 2005. SEPALLATA gene diversification: brave new whorls. Trends Plant Sci.. 10, 427-435.
|
| [29] |
McSteen, P., Laudencia-Chingcuanco, D., Colasanti, J., 2000. A floret by any other name: control of meristem identity in maize. Trends Plant Sci.. 5, 61-66.
|
| [30] |
Muramatsu, M., 1963. Dosage effect of the spelta gene q of hexaploid wheat. Genetics, 48, 469.
|
| [31] |
Muramatsu, M., 1986. The vulgare super gene, Q: its universality in durum wheat and its phenotypic effects in tetraploid and hexaploid wheats. Can. J. Genet. Cytol. 28, 30-41.
|
| [32] |
Poethig, R.S., 1990. Phase change and the regulation of shoot morphogenesis in plants. Science 250, 923-930.
|
| [33] |
Sanchez-Leon, S., Gil-Humanes, J., Ozuna, C.V., Gimenez, M.I.A.J., Sousa, C., Voytas, D.F., Barro, F., 2018. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 16, 902-910.
|
| [34] |
Sakuma, S., Golan, G., Guo, Z., Ogawa, T., Tagiri, A., Sugimoto, K., Bernhardt, N., Brassac, J., Mascher, M., Hensel, G., Ohnishi, S., Jinno, H., Yamashita, Y., Ayalon, I., Peleg, Z., Schnurbusch, T., Komatsuda, T., 2019. Unleashing floret fertility in wheat through the mutation of a homeobox gene. Proc. Natl. Acad. Sci. U. S. A. 116, 5182-5187.
|
| [35] |
Simons, K.J., Fellers, J.P., Trick, H.N., Zhang, Z., Tai, Y.S., Gill, B.S., Faris, J.D., 2006. Molecular characterization of the major wheat domestication gene Q. Genetics 172, 547-555.
|
| [36] |
Song, G., Sun, G., Kong, X., Jia, M., Wang, K., Ye, X., Zhou, Y., Geng, S., Mao, L., Li, A., 2019. The soft glumes of common wheat are sterile-lemmas as determined by the domestication gene Q. Crop J. 7, 113-117.
|
| [37] |
Tang, M., Li, G., Chen, M., 2007. The phylogeny and expression pattern of APETALA2-like genes in rice. J. Genet. Genomics 34, 930-938.
|
| [38] |
Varkonyi-Gasic, E., Lough, R.H., Moss, S.M., Wu, R., Hellens, R.P., 2012. Kiwifruit floral gene APETALA2 is alternatively spliced and accumulates in aberrant indeterminate flowers in the absence of miR172. Plant Mol. Biol. 78, 417-429.
|
| [39] |
Wang, K., Liu, H., Du, L.m Ye, X., 2017. Generation of marker-free transgenic hexaploid wheat via an Agrobacterium-mediated co-transformation strategy in commercial Chinese wheat varieties. Plant Biotechnol. J. 15, 614-623.
|
| [40] |
Wang, M., Wang, S.B., Liang, Z., Shi, W.M., Gao, C.X., Xia, G.M., 2018. From genetic stock to genome editing: gene exploitation in wheat. Trends Biotechnol.. 36, 160-172.
|
| [41] |
Wang, Y., Cheng, X., Shan, Q., Zhang, Y., Liu, J., Gao, C., Qiu, J., 2014. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 32, 947.
|
| [42] |
Wang, Y., Li, J., 2008. Molecular basis of plant architecture. Annu. Rev. Plant Biol. 59, 253-279.
|
| [43] |
Whipple, C.J., Ciceri, P., Padilla, C.M., Ambrose, B.A., Bandong, S.L., Schmidt, R.J., 2004. Conservation of B-class floral homeotic gene function between maize and Arabidopsis. Development 131, 6083-6091.
|
| [44] |
Wu, D., Liang, W., Zhu, W., Chen, M., Ferrandiz, C., Burton, R.A., Dreni, L., Zhang, D., 2018. Loss of LOFSEP transcription factor function converts spikelet to leaf-like structures in rice. Plant Physiol.. 176, 1646-1664.
|
| [45] |
Yamaguchi, T., Lee, D.Y., Miyao, A., Hirochika, H., An, G., Hirano, H., 2006. Functional diversification of the two C-class MADS box genes OSMADS3 and OSMADS58 in Oryza sativa. Plant Cell 18, 15-28.
|
| [46] |
Yoshida, A., Ohmori, Y., Kitano, H., Taguchi-Shiobara, F., Hirano, H., 2012. ABERRANT SPIKELET AND PANICLE1, encoding a TOPLESS-related transcriptional co-repressor, is involved in the regulation of meristem fate in rice. Plant J.. 70, 327-339.
|
| [47] |
Zhang, K., Liu, J., Zhang, Y., Yang, Z., Gao, C., 2015. Biolistic genetic transformation of a wide range of Chinese elite wheat (Triticum aestivum L.) varieties. J. Genet. Genomics 42, 39-42.
|
| [48] |
Zhang, S., Zhang, R., Gao, J., Gu, T., Song, G., Li, W., Li, D., Li, Y., Li, G., 2019a. Highly efficient and heritable targeted mutagenesis in wheat via the Agrobacterium tumefaciens-mediated CRISPR/Cas9 system. Int. J. Mol. Sci. 20, 4257.
|
| [49] |
Zhang, Y., Li, D., Zhang, D., Zhao, X., Cao, X., Dong, L., Liu, J., Chen, K., Zhang, H., Gao, C., Wang, D., 2018. Analysis of the functions of TaGW2 homoeologs in wheat grain weight and protein content traits. Plant J.. 94, 857-866.
|
| [50] |
Zhang, Y.W., Bai, Y., Wu, G.H., Zou, S.H., Chen, Y.F., Gao, C.X, Tang, D.Z., 2017. Simultaneous modification of three homoeologs of TaEDR1 by genome editing enhances powdery mildew resistance in wheat. Plant J.. 91, 714-724.
|
| [51] |
Zhang, Z., Belcram, H., Gornicki, P., Charles, M., Just, J., Huneau, C., Magdelenat, G., Couloux, A., Samain, S., Gill, B., Rasmussen, J., Barbe, V., Faris, J., Chalhoub, B., 2011. Duplication and partitioning in evolution and function of homoeologous Q loci governing domestication characters in polyploid wheat. Proc. Natl. Acad. Sci. U. S. A. 108, 18737-18742.
|
| [52] |
Zhang, Z., Hua, L., Gupta, A., Tricoli, D., Edwards, K.J., Yang, B., Li, W., 2019b. Development of an Agrobacterium-delivered CRISPR/Cas9 system for wheat genome editing. Plant Biotechnol. J. 17, 1623-1635.
|
| [53] |
Zhang, Z., Li, A., Song, G., Geng, S., Gill, B.S., Faris, J.D., Mao, L., 2020. Comprehensive analysis of Q gene near isogenic lines reveals key molecular pathways for wheat domestication and improvement. Plant J.. 102, 299-310.
|
| [54] |
Zhao, K., Xiao, J., Liu, Y., Chen, S., Yuan, C., Cao, A., You, F., Yang, D., An, S., Wang, H., Wang, X., 2018. Rht23 (5Dq′) likely encodes a Q homeologue with pleiotropic effects on plant height and spike compactness. Theor. Appl. Genet. 131, 1825-1834.
|
| [55] |
Zhu, J., Song, N., Sun, S., Yang, W., Zhao, H., Song, W., Lai, J., 2016. Efficiency and inheritance of targeted mutagenesis in maize using CRISPR-Cas9. J. Genet. Genomics 43, 25-36.
|
| [56] |
Zhu, Q.H., Helliwell, C.A., 2011. Regulation of flowering time and floral patterning by miR172. J. Exp. Bot. 62, 487-495.
|

Figures (7) / Tables (2)





Supported by: Beijing Renhe Information Technology Co. Ltd




 DownLoad:
DownLoad: